研究紹介
1. フラボノイド配糖体アピインの生合成と機能
特化代謝産物
動けない植物は環境適応性を高める仕組みを持っています。その一つは特化代謝産物(二次代謝産物) を生産することです。特化代謝産物は多様で数十万種類あります。例えば、アントシアニンは花色をつけ、花粉媒介昆虫を引き寄せ、レスベラトロールは病原菌などの外敵に対抗しています。特化代謝産物はヒトにも作用します。コーヒーやお茶に含まれるカフェインは有名です。ケルセチンやカテキンが脂肪代謝を活性化させます。それらを配合しているサントリー特茶、花王ヘルシアなどの商品がヒットしています。イチイの樹皮から採られるパクリタキセルはがん化学療法に用いられています。
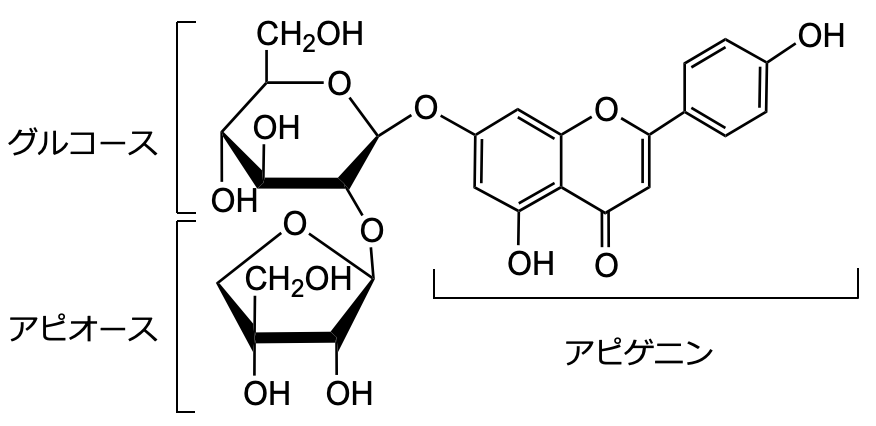
パセリが多く生産するフラボノイド配糖体アピイン
パセリ(下図)はアピイン(下図)というフラボノイド配糖体(ポリフェノールの一種)を生産しています。アピオースという変わった糖が含まれています。パセリは害虫が少なくキアゲハしか産卵しないので、昆虫忌避物質があるとされています。古代ヨーロッパではパセリは神経疾患や不安を和らげるものとして用いられ、現代でもハーブの一種とされています。カモミールもアピインを生産します。パセリは植物重量の2%分もアピインを生産しています。このため、アピインが昆虫忌避物質であったり、抗不安作用物質であると考えられ始めています。
アピインの生合成と機能
アピインの機能を明らかにするためにはアピインを生合成する酵素遺伝子を同定しなければなりません。石水研では、アピインを生合成するアピオース転移酵素の遺伝子を世界に先駆けて同定しました(Fujimori et al., 2019 ; Yamashita et al., 2023 )。この遺伝子が同定されたことで、アピイン生合成に関わる10種類以上の酵素のすべてが同定されたことになりました。これにより、アピインを生産することやアピインの機能を解析できることになり、それらの研究を進めています 。また、複数の酵素の複合体メタボロンの解析も進めています。
2.植物細胞壁ペクチン生合成の分子機構
植物細胞壁多糖成分ペクチン
植物細胞壁は、地球上で最も多く存在する生物資源で、エネルギー源としても注目されています。セルロース、ペクチン、ヘミセルロースなどの多糖から構成されています。このうち、ペクチンは植物成長・細胞接着・植物のしなやかさの調整に関わるとされています。ジャムやゲル化剤として広く食品産業に利用されています。フルーチェ(ハウス食品)やコバラサポート(大正製薬)の機能成分や漢方薬の有効成分もペクチンです。このようにペクチンは、植物にとっても人にとってもなくてはならない物質です。
ペクチンは、知られている糖鎖の中で最も複雑な構造をしています。構造が複雑すぎて、どのようにペクチンが合成されるのか、未だに明確になっていません。石水研では、ペクチン合成酵素を見つける研究に取り組んでいます。ペクチン合成の仕組みが明らかになると、ペクチンの機能や利用の研究が進みます。例えば、植物のしなやかさの調整の仕組みを解き明かしたり、成長しやすい植物(食糧増収に貢献)・分解しやすい多糖をもつ植物(バイオエネルギーにしやすい植物)を作成したり、新しい食材・医薬品(機能化ゲル化剤、漢方薬成分など)を作り出すことに活かされます。
ペクチンを合成する糖転移酵素
ペクチンは、糖転移酵素によって一つ一つ糖をつなげて合成されます。ペクチンは12種類の糖と約30種類の結合様式から構成されているため、約30種類の糖転移酵素がペクチン合成に関わると考えられています。当研究室では、糖転移酵素の基質化合物を調製し、酵素の活性測定法を構築しています(Akita et al., 2002 ; Ishimizu et al., 2005 ; Ohashi et al., 2006 ; Uehara., 2017 ; Matsumoto et al., 2019 ; Fujimori et al. 2019 )。この方法を用いて、ペクチン主鎖を合成する糖転移酵素遺伝子を世界に先駆けて発見しました(Takenaka et al., 2018 )(下図)。糖転移酵素ファミリー106(GT106) という新しい酵素のファミリーの発見にもつながりました。しかし、ペクチン合成に関わる多くの酵素が未発見のままです。一つ一つ発見し、ペクチン合成の全貌を明らかにすることを目指しています。
ペクチンの構造は複雑であるにも関わらず、種を越えて一定です。そのため、ペクチン合成に関わる複数の酵素が連携した生合成の機構があると考えられています。石水研では、これまでに開発した膜タンパク質取扱技術(Yasui et al., 2010 ; Uegaki et al., 2010 ; Arakawa et al., 2011 )を生かして、膜タンパク質であるペクチン生合成酵素の複合体を解析することにチャレンジしています。
ペクチン合成と植物進化
ペクチンは、陸上植物にはあり、水生植物にはありません。石水研で発見したペクチン合成酵素が陸上植物にはあり水生植物にはなかった(下図)ことから、進化の過程で植物が5億年前に陸上化したのは、ペクチン合成酵素の獲得によって、重力に逆らって植物が垂直に立つ細胞強度を得たためと考えられました(Takenaka et al., 2018 )。陸上化した原始植物と考えられる車軸藻やコケ植物を用いて、ペクチンの機能研究も展開しています(Wachananawat et al., 2020 )
3.植物糖タンパク質糖鎖の分解の分子機構
特徴的な植物の糖タンパク質糖鎖
真核生物のタンパク質の多くは糖鎖修飾を受けた糖タンパク質です。糖鎖は、物性的にはタンパク質の安定化、機能的にはタンパク質輸送や分解に関わることがわかっています。植物の糖タンパク質糖鎖は、哺乳動物のものとは違い、構造が特徴的です。マンノース5残基が結合しているハイマンノース型糖鎖やフコース・キシロース残基を含む植物コンプレックス型糖鎖が多く存在します。石水研では、ジアセチルキトビオース構造が、植物に特徴的にあることを発見しています(Ishimizu et al., 1996 ; Ishimizu et al., 1999 )。
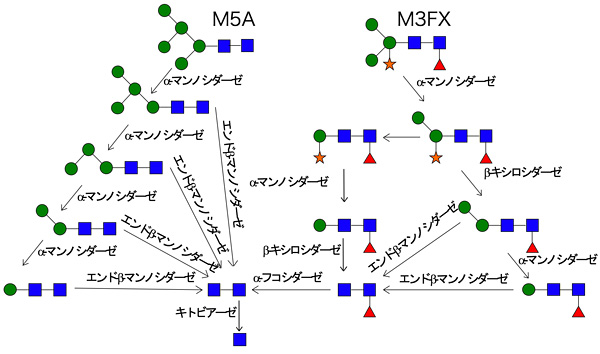
植物糖タンパク質糖鎖の分解経路
植物に特徴的な糖鎖の生成過程を研究することで、植物特異的糖質分解酵素を見つけました(Ishimizu et al., 2004 ; Kato et al., 2018 )。これらの糖質分解酵素の性質を明らかにすることで、植物特有の糖鎖分解経路があることを見つけました(下図)(石水, 2011 ; Kato et al., 2018 )。糖鎖分解に関わる複数の酵素が同定されたので、協同的に糖鎖分解を行うシステムを対象とした研究を進めることができるようになります。
4. 新規酵素の発見
石水研の得意分野は酵素研究です。糖鎖に作用する酵素は糖質関連酵素(Carbohydrate-Active Enzymes; CAZymes )と呼ばれています。ペクチン、フラボノイド配糖体、糖タンパク質糖鎖に作用するCAZymesを発見しています。ペクチン合成する酵素は、まだ不明なものが数多く残されており、それらの酵素を発見する研究を続けています。酵素は基質特異性によりEC番号に分類され、これまでに6754種類(2023年9月13日現在)の酵素が登録されています。このうち、石水研で3種類の酵素を発見しています。
糖質関連酵素はアミノ酸配列によっても分類され、CAZyデータベース でファミリー分けされています。糖質加水分解酵素(GH) は185ファミリー、糖転移酵素(GT) は116ファミリーに分けられています。このうち、石水研でGT106ファミリーを発見しています。
酵素遺伝子を同定する研究にも取り組んでいて、ペクチンRG-Iラムノース転移酵素遺伝子(Takenaka et al., 2018 )、フラボノイド配糖体アピいん生合成アピオース転移酵素(Yamashita et al., 2023 )を同定しています。
5. イネの倒伏に関与する遺伝子の単離とその機能解析
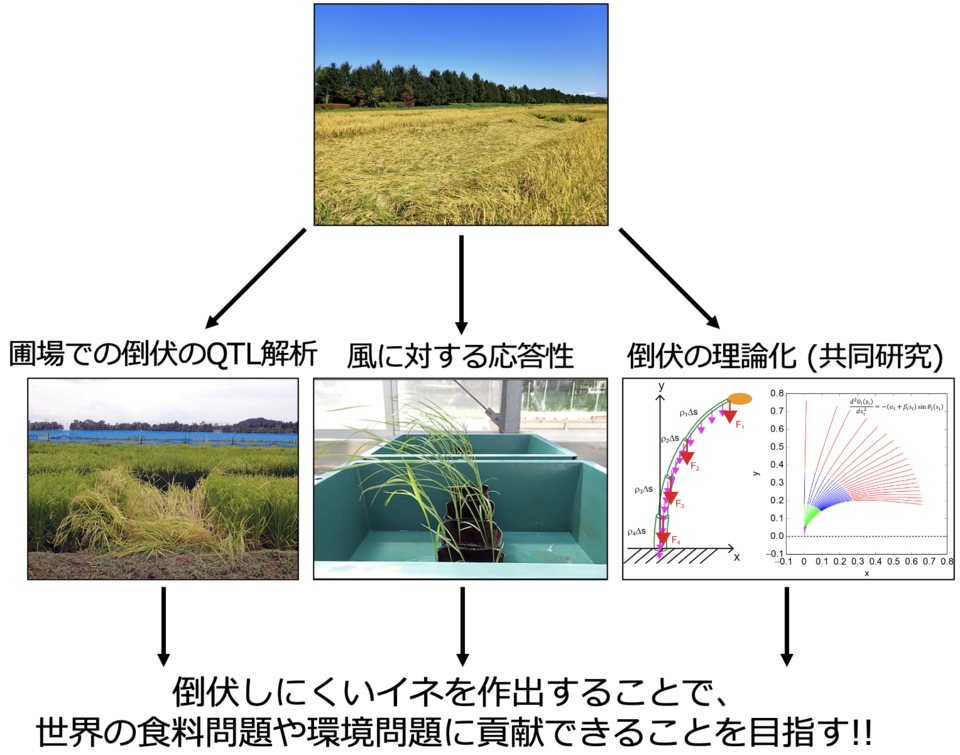
世界の主要三大穀物の一つであるイネは、病気や高温、低温、塩害だけでなく、台風などの暴風雨によって引き起こされる倒伏によっても、収量や品質の低下が引き起こされます。そのため、倒伏に強いイネを作出することは、世界の食料問題を解決するためにも非常に重要です。しかしながら、イネがどのように暴風雨に抵抗しているのか、イネの倒伏にはどの組織 (根、稈のどの節間、穂、葉など) が重要か、背丈など形の違うイネの倒伏に対する抵抗性 (倒伏のしにくさ) をどのように数値化するかなど、多くのことがまだ解明されていません。当研究室では、圃場における倒伏のQTL解析や風に対するイネの応答機構、数理解析による倒伏の理論化 (秋田県立大学、山梨大学、京都大学との共同研究) を行うことで、これらの解明を目指しています。それにより、我々の研究成果が、世界の食料問題や環境問題に貢献できることを期待しています。
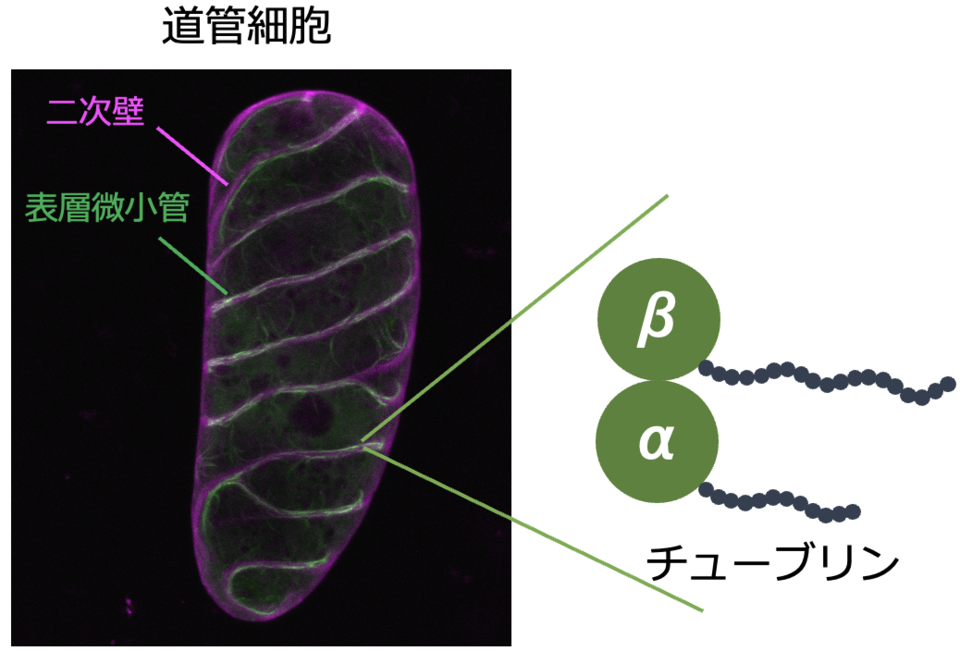
6. 道管細胞分化における二次細胞壁パターン形成制御機構の解明
道管は水や無機塩類の輸送、植物体の支持を担う植物にとって重要な組織です。維管束木部組織を構成する道管細胞や繊維細胞は一次細胞壁と細胞膜の間に二次細胞壁(二次壁)を形成します。二次壁はらせん状等の特徴的なパターンで沈着することで細胞に機械的な強度や生化学的な特性を付与します。また、二次壁が沈着しない領域は水の通り道となることからも、二次壁パターンは通水機能発現において重要な構造です。この二次壁を構成する成分の沈着は細胞膜下の表層微小菅の配向に沿ってみられます。表層微小管は二次壁パターンだけでなく、全ての細胞の形態形成や細胞分裂においても重要な役割を果たしています。それらの機能の違いには微小菅を構成するタンパク質や結合タンパク質が関与しています。私たちは、微小菅構成タンパク質であるチューブリンに着目し、二次壁パターン形成機構に特異的な表層微小菅制御機構解明を目指しています。